
Nederlandse vertaling van een artikel uit Elsevier Science (USA).
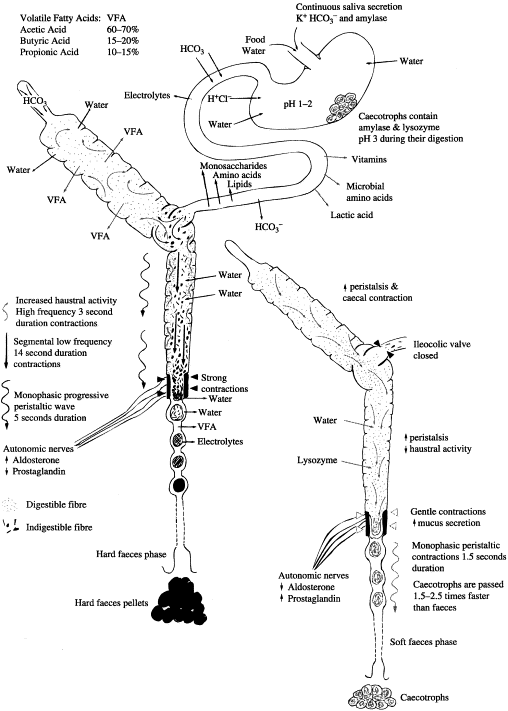
Het konijn is een herbivoor (planteneter), of meer specifiek een folivoor (bladeter), die uitermate geschikt is om te overleven op een dieet van groene vegetatie. Doordat het konijn klein is, heeft het een hoge stofwisseling, waardoor het niet kan overleven op een dieet met een lage energiedichtheid. Het gevolg hiervan is dat het konijn een zeer goede prooi is, die op zijn beurt zeer wendbaar en atletisch dient te zijn om roofdieren te snel af te zijn. Om met deze problemen om te kunnen gaan, heeft het konijn een totaal verschillend spijsverteringskanaal ontwikkeld dan andere planteneters zoals het paard (verteert voedsel in de dikke darm) en andere knaagdieren (verteren het voedsel in de maag). Het konijn heeft een spijsverteringsstelsel dat: (1) een hoge voedselinname toelaat (en daardoor hoge energie-inname en inname van eiwitten), (2) makkelijk verteerbare voedingsbestanddelen scheidt uit het voedsel en (3) zeer snel de niet- of moeilijk verteerbare bestanddelen uitscheidt en deze dus niet hoeft mee te dragen. Dit stelsel maakt ook een groot absorptief oppervlakte van de dikke darm overbodig, doordat de voedingsstoffen gescheiden worden van de faeces, dat nieuwe inname van bacteriën en de bijproducten toestaat in de dunne darm. Gegeven het feit dat het systeem is ontworpen om niet-verteerbare producten zoals vezels snel uit te kunnen scheiden contrasteert ironisch genoeg met het feit dat het dieet van een konijn is opgebouwd rond producten die juist veel vezels bevatten. Het ontbreken van vezels aan het dieet van het konijn is de voornaamste oorzaak van verstoring van het spijsverteringsstelsel. Het doel van het artikel is om de tegenwoordige kennis van het spijsverteringsstelsel van het konijn onder de loep te nemen en markeert een aantal gevallen waarbij afbraak van de normale fysiologie van het stelsel tot ziekte leidt. Figuur 1 (onderaan dit artikel) is een schematisch diagram ter referentie van het spijsverteringsstelsel van het konijn en figuur 2 is een weergave van de activiteit van het verteringsstelsel.
De mondholte
Inname van voedsel
Wilde konijnen eten selectief jonge enten. Om deze te pakken te krijgen gebruiken ze hun snijtanden, die de vorm van een vijzel hebben, om deze korte stukjes vegetatie af te snijden. Deze manier van voedsel vergaring wordt mogelijk gemaakt door het diastema, rostraal gepositioneerde snijtanden en de gespleten bovenlip (hazenlip). De localisatie van het voedsel vindt plaats door middel van gevoelige trilharen op de lippen, doordat de positie van de ogen van konijnen voorkomt dat ze voorwerpen kunnen zien, die zich recht voor de mond bevinden. De lippen zelf zijn uiterst beweegbaar en in gevangenschap (waar het dieet voornamelijk bestaat uit hooi met lange stam, stukken groente en gepelleteerd of ander voedsel) zijn de snijtanden voor het grootste deel overvloedig. Daarom kunnen konijnen in gevangenschap gemakkelijk omgaan met verwijdering van de snijtanden.
Kauwen
Na inname van het voedsel, wordt het fijngemalen door de zogenaamde wangtanden (voorkiezen en kiezen). De tanden van elk compartiment zijn zeer dicht tegen elkaar geplaatst en functioneren meer als een groot oppervlak dan als afzonderlijke tanden. De marges van elke tand en de rand, die loopt van de lippen tot de kaak, bestaan uit enamel. De valleien tussen de structuren bestaan uit het zachtere dentine.
De voortdurende slijtage door het voedsel en het malen van de tanden zorgt ervoor dat de vlakke oppervlakte van de tanden behouden blijft en dat er een uitstekend oppervlak ontstaat om het voedsel te vermalen. De bewegingen van de kaken en tanden gedurende het kauwen zijn reeds beschreven en in beeld gebracht, alsmede de gecoördineerde bewegingen van de tong gedurende het proces. De handelingen gedurende het kauwen zijn onderverdeeld in drie verschillende types. Type I acties zijn betrokken bij het afsnijden van plakjes door de snijtanden bij het grijpen van voedsel. Type II acties beslaan het grootste deel van het kauwproces en bestaan uit het kauwen en fijnmalen van voedsel en de afbraak van de lange vezels van het hooi in kleinere delen. Slechts een zijde van de mond wordt tegelijk gebruikt tijdens dit kauwproces en de onderkaak beweegt altijd naar het midden op het moment dat druk wordt uitgeoefend op het voedsel. Type III acties beslaan het vormen van een brok voedsel en dus het voorbereiden van het voedsel om door te slikken. Tot wel 120 kaakbewegingen per minuut zijn gerapporteerd bij het verwerken van vers voedsel. Wanneer blindedarmkeutels worden gegeten vinden type II acties niet plaats, maar worden deze in hun geheel doorgeslikt.
Speekselsecretie
Het konijn heeft vier grotere paren speekselklieren: onder de oren, in de onderkaak, onder de tong en in de wangen. De twee enzymen amylase en galactosidase worden geproduceerd in het speeksel, dat voortdurend wordt afgegeven vanuit de klieren in de onderkaak en wanneer voedsel wordt ingenomen ook door de andere speekselklieren 9]. Lipase en ureum (de voornaamste enzymen in menselijk speeksel en dat van hoefdieren) zijn slechts sporadisch te vinden in het speeksel van konijnen. Kalium en bicarbonaat-ionen zijn ook belangrijke bestanddelen van het speeksel van konijnen.
De slokdarm
De slokdarm dient als een transportkanaal van de mondholte naar de maag. De structuur en functie van de slokdarm wijken in de konijnen nauwelijks af van andere niet-hoefdieren en hebben (bijna) geen effect op de spijsvertering. Verdere beschrijving van de slokdarm is overbodig.
De maag
Anatomie van de maag
De maag van een konijn is een orgaan in de vorm van een zak met een dunne wand. De maag beslaat ongeveer 15% van het totale volume van het spijsverteringskanaal. De maag bevat een zeer goed ontwikkelde maagzak die ervoor zorgt dat een konijn kan spugen. De maagzak heeft ook een dunne wand, geen klieren en immobiel. Wanneer voedsel in de maagzak terechtkomt is dat meestal eerder het gevolg van bewegingen van de darmen en van het konijn zelf dan van de bewegingen van de maag. Normaal gesproken is de maag van een konijn nooit leeg. Het is zelfs aangetoond dat de maag van een konijn na een periode van vasten van 24 uur nog steeds voor 50% gevuld was, normaal gesproken met een voedselmassa en haren omgeven door vloeistoffen.
Van haarballen werd gedacht dat deze het gevolg waren van de inname van vacht, maar worden nu in het algemeen beschouwd als het gevolg van een verminderde beweeglijkheid van de maag, vaak ten gevolge van inactiviteit van het konijn of op de tweede plaats door verminderde darmperistaltiek.
De maagzak is het belangrijkste deel van de maag voor de uitscheiding van verteringssappen en bevat twee soorten cellen. Parietale cellen scheiden zoutzuur en een intrinsieke factor uit en peptische (eiwit-) cellen zorgen voor de secretie van pepsinogeen, de voorloper van pepsine. De pylorus (sluitspier van de maag) heeft een veel dikkere wand dan de overige compartimenten van de maag.
Fysiologie van de maag van zuigelingen en veranderingen na de overstap naar gewoon voedsel
Zuigelingen hebben een pH (zuurgraad) in de maag van 5 – 6,5. De ingenomen melk vormt een halfvaste massa in de maag, die geleidelijk wordt doorgegeven naar de dunne darm gedurende de 23,5 uur tussen de voedingen. De vorming van de halvaste massa is het gevolg van de actie van een rennine-achtig enzym. In de meeste andere dieren zou deze ophoping van voedsel zorgen voor verhoogde groei van bacteriën. Zuigelingen hebben op deze leeftijd echter een bescherming tegen bacteriële infectie door de aanwezigheid van ‘maagolie’ of ‘melkolie’- een product van vetzuren (octaan- of decaan-) met antimicrobiële eigenschappen, gevormd door de enzymen van de zuigeling uit de moedermelk. Zelfs wanneer de bacteriën wel zouden groeien, hebben zuigelingen nog passief verkregen antilichamen van de moeder (uit de placenta en de eerste zuigelingvoeding) tegen bacteriën en andere microben. Al deze factoren zorgen ervoor dat het spijsverteringskanaal van een zuigeling bijna steriel blijft. Jonge konijnen die met de hand melk krijgen of de melk van een ander dierenras krijgen, kunnen geen antimicrobiële substantie vormen, hevige bacteriële enteritis komt ook veelvuldig voor bij konijnen die met de fles gevoed worden. Totdat de zuigelingen 10 dagen oud zijn, zijn ze volledig afhankelijk van melk. Naarmate de konijnen ouder worden, beginnen ze blindedarmkeutels van de moeder te eten. Doordat deze keutels vrij lang intact blijven in de maag binnen een slijmlaag, blijven de bacteriën in de keutels lang genoeg beschermd tegen de maagolie om uiteindelijk het laatste deel van de darm te kunnen koloniseren. Deze bacteriën heeft het konijn nodig om het voedsel goed te kunnen verteren. Vanaf 15 dagen beginnen jonge konijnen met de consumptie van wat vast voedsel. Na 20 dagen vormt vast voedsel het grootste deel van het dieet en beginnen de konijntjes met het leggen van keutels en na 30 dagen is de inname van melk minimaal en de vorming van keutels volledig ontwikkeld. Gedurende dezelfde periode neemt de vorming van maagolie af. De pH van de maag neemt af naar het niveau van volwassen konijnen (1-2), wat een nieuwe effectieve barrière opwerpt tegen bacteriële infectie van de maag en darmen. De effectiviteit van de bescherming van het konijn tegen bacteriëlendeldar infectie gedurende de overgang van zuigeling naar niet-zuigeling hangt af van de synchronisatie van de overgang van het ene mechanisme naar het andere. Het overgrote deel van darmziekte incidentie ( bijvoorbeeld coliforme infecties, coccidiose, mucoid enteropathy syndroom en Rotavirus-gerelateerde diarree) vindt dus plaats gedurende de periode direct na de overstap op vast voedsel.
Fysiologie van de maag na de overstap op vast voedsel
In het volwassen konijn worden grote hoeveelheden water en zuur uitgescheiden in de holte van de maagzak. De pH van de maag van een volwassen konijn wordt gedurende de inname van voedsel constant gehouden tussen 1 en 2, dat ervoor zorgt dat het grootste deel van de microben in het voedsel kapot maakt en dus resulteert in een bijna steriele maag en dunne darm. De duur van de passage van voedsel door de maag wordt geschat op 3-6 uur. Hydrolyse van eiwitten begint in de maag, met het pepsine-HCl complex. Een belangrijke uitzondering is de vertering van het slijm rond de blindedarmkeutels. Deze keutels worden niet gemalen door de tanden en blijven intact in de maag, beschermd door de slijmlaag voor minstens 6-8 uur na inname. Gedurende deze periode zijn de bestanddelen beschermd tegen de werking van de pH van de maag en microben blijven groeien, wat leidt tot de vorming van melkzuur in de maag. De pH van de maag stijgt gedurende de periode naar 3 door de bufferende werking van het lactaat dat aangemaakt wordt door de bacteriën in de blindedarmkeutels. Dit proces heeft geleid tot het idee dat blindedarmkeutels probiotisch effecten hebben in de pathologie van de dikke darm (bijvoorbeeld antibiotisch-gerelateerde dysenterobiose, enteritis). Er zijn echter twee belangrijke tegenstrijdigheden.
Op de eerste plaats is het haast onmogelijk om de blindedarmkeutels in hun geheel door te slikken zonder de slijmlaag te breken. Ten tweede is het onmogelijk voor de keutels om in hun geheel de maag te passeren naar de dunne darm totdat de slijmlaag verbroken is, zodat de bacteriën blootstaan aan het normale verteringsproces en niet overleven om de darmen te koloniseren.
De dunne darm
Anatomie en beweeglijkheid van de dunne darm
De beweeglijkheid van de dunne darm kan worden onderverdeeld in een aantal verschillende processen. Segmentatie is het proces waarbij de inhoud van de darmen wordt vermengd door de statische peristaltische samentrekkingen van de darmwand en is zeer belangrijk in de twaalfvingerige darm van het konijn. Een ander proces in de beweeglijkheid van de darm is de peristaltiek. Dit proces bevat een ring van samentrekking van de darmwand die geleidelijk beweegt langs de darm, normaal gesproken in de richting vanaf de mond. De regulatie van de peristaltiek omvat een aantal gastro-intestinale hormonen en enzymen, onder andere cholecystokinine, somatostatine vaso-actief peptide en de ‘substantie P’. De passage van voedsel door de dunne darm gaat snel in verhouding tot andere planteneters. Peristaltiek vindt langzaam plaats, enkele 10-15 minuten en veranderen niet. De tijd die nodig is voor de spijsvertering is geschat op 10-20 minuten in de jejunum (onderdeel van de dunne darm) en 30-60 minuten in de ileum (onderdeel van de dunne darm). De beweeglijkheid van de dunne darm is bij het konijn, net als bij de mens en in tegenstelling tot de meeste andere diersoorten, gedeeltelijk gereguleerd door motiline, een eiwit dat wordt uitgescheiden door enterochromaffine cellen van de twaalfvingerige darm (onderdeel van de dunne darm) en de jejunum. Motiline stimuleert de samentrekking van gladde spiercellen. De afgifte van motiline wordt gestimuleerd door de aanwezigheid van vetten en geremd door de aanwezigheid van koolhydraten (suikers) in de darminhoud. Macrolide antibiotica kunnen in sommige gevallen ook optreden als stimulatoren van motiline en peristaltiek. De activiteit van motilin neemt af naarmate het voedsel in het achterste deel van de dunne darm aankomt, maar neemt weer toe in de dikke darm en het rectum. Aan het eind van het verre uiteinde van de ileum, aan de achterkant van de dikke darm gelegen, is een ronde gespierde ampul gesitueerd, die ook wel sacculus rotundus genoemd wordt (figuur 1). Deze structuur schijnt een immunologische functie te hebben en is allen aanwezig bij knaagdieren. Dit is een van de belangrijkste plaatsen voor het tegenhouden van vreemde lichamen uit de darmen. Een klep tussen ileum en sacculus rotundus voorkomt het terugstromen van vloeistoffen in de ileum en zorgt dat de voedselbrij via de sacculus rotundus naar de blindedarm gaat.
Secretie uit de alvleesklier en de lever
Zoals verwacht bij dieren met een constant actief verteringssysteem en lage inname van koolhydraten en eiwitten is de alvleesklier maar heel klein. Het is diffuus en vaak zeer moeilijk te vinden in het mesenterisch vetweefsel tussen de dikke darm, de maag en de twaalfvingerige darm. Het belangrijkste kanaal vanuit de alvleesklier eindigt bij het einde van de twaalfvingerige darm, een eind verwijderd van de galbuis.
Hoewel ligatie van de kanalen van de alvleesklier leidt tot verwijding van de kleinere kanaaltjes, komen enzymen uit de alvleesklier toch voor in de holte van de ileum, wat duidt op de aanwezigheid van andere zeer kleine kanaaltjes. Trypsine, chymotrypsine en carboxypeptidases worden geproduceerd in de alvleesklier en afgegeven in de darm. Samen met aminopeptidases uit de darm voltooien ze de vertering van eiwitten. Verder worden allerlei vormen van lipases gevormd. De alvleesklier is een belangrijke bron voor bicarbonaat-ionen die de zure voedselbrij, die vanuit de maag in de darm terechtkomt, neutraliseren.
Vanuit de lever lopen galbuizen naar de galblaas, die net iets van de sluitspier van de maag verwijderd ligt. Een konijn maakt ongeveer 100-150 mL gal aan per kg lichaamsgewicht per dag, onafhankelijk van de stimulatie door secretine – zeven keer meer dan een hond.
Galzouten, zoals cholinezuur en chenodeoxy-cholinezuur, worden geproduceerd door de lever en afgegeven in de dunned arm, waar een deel ervan wordt omgezet in deoxy-cholinezuur door microbiële activiteit.
Gal is belangrijk als emulgator om vetzuren en olieachtige stoffen te verdelen over kleine micellen, die de opname van vetten en vetoplosbare vitamines toelaten in de dunne darm. De andere functionele bestanddelen van gal zijn de pigmenten. Biliverdine is het afbraakproduct van hemoglobine en wordt in de meeste dieren door het enzym biliverdine-reductase afgebroken tot bilirubine, voordat ze worden uitgescheiden in gal.
De activiteit van biliverdine-reductase is laag bij het konijn, zelfs 60 maal lager dan in ratten en 63% van de pigmenten in konijnen is niet-omgezette biliverdine.
Opname en uitscheiding in de dunne darm
Vertering en opname in de dunne darm van het konijn is gelijk aan andere dieren. Bicarbonaat-ionen worden uitgescheiden in de twaalfvingerige darm om de zuurgraad van de voedselbrij uit de maag te verminderen. Het grootste deel van de vertering van suikers en simpele eiwitten vindt plaats in de twaalfvingerige darm en de jejunum en de producten van deze afbraak (monosacchariden en aminozuren) worden opgenomen door het membraan van de darm. Hiertoe behoren ook de vertering en opname van aminozuren, vrije vetzuren, vitamines en verteerde micro-organismen uit de darmkeutels. Deze vertering wordt gemedieerd door toevoeging van lysozym aan de keutels wanneer ze de dikke darm passeren.
Afbraak van de microben maakt ook microbiële enzymen vrij, voornamelijk amylase, dat de verteringsprocessen in het konijn verbetert. De ileum speelt tenslotte een belangrijke rol bij de regulatie en het hergebruik van elektrolyten, die zijn vrij gekomen in de maag en het eerste deel van de dunne darm door de heropname van bicarbonaat-ionen.
De dikke darm
Blinde darm en wormvormig aanhangsel: anatomie, microbiologie, en biochemie
De blindedarm van het konijn is proportioneel het grootst ten opzichte van ieder ander zoogdier. Het beslaat tweemaal de lengte van de buikholte en ongeveer 40-60% van het totale volume van het spijsverteringskanaal]. Het is een zakvormig orgaan dat in vier delen (gyri) is gevouwen. De eerste gyrus loopt van de navelstreek in de richting van de schedel naar rechts, dwars over de bodem van de buikholte. Vervolgens buigt het in de richting van de staart en de tweede gyrus buigt weer verder naar de staart en loopt parallel in tegengestelde richting van de eerste gyrus naar links dwars over de bodem van de buikholte. De derde gyrus loopt weer in de richting van het hoofd en loopt langs de linker flank van de buik en loopt wederom parallel aan de vorige gyrus, deze keer dichter naar het hoofd toe en gescheiden door het opwaartse deel van de dikke darm. Een lange spiraal vormt de laatste vouw van de cecocolische ampul bij de verbinding met de dikke darm, met 20-30 bochten. De eerste drie gyri hebben een dunne doorschijnende wand. De vierde gyrus (het wormvormig aanhangsel) is een 12,5 cm lange, doodlopende buis, die eindigt aan de linker flank, dorsaal (naar de rug toe) van het eerste deel van de blinde darm en heeft dikke wanden met veel lymfe weefsel.
Het aanhangsel scheidt bicarbonaat-ionen uit in de blindedarm, die als buffer optreden voor de vrije vetzuren die geproduceerd zijn bij de vertering. Konijnen die een dieet krijgen met weinig vezels en veel suikers, ontwikkelen een vergrote appendix. Dit is gebruikt als bewijs dat een verhoogde uitscheidende functie van de appendix nodig is om de producten van de suikerstofwisseling tegen te gaan. Het kan echter ook als bewijs worden gebruikt voor de noodzaak van het lymphoide weefsel door veranderingen in de bacteriekolonies in de darmen door verhoogde suikerinname in zulke konijnen.
Water, uitgescheiden door de appendix en de dikke darm, wordt constant toegevoegd aan de darminhoud, waarna het weer wordt geabsobeerd in de darmwand. De darminhoud blijft halfzacht. De normale pH van de blindedarm van een konijn varieert binnen de verteringscyclus (zie onder). De pH in de middag (meest zure periode) is normaal gesproken hoger in volwassen konijnen (5,9-6,8) dan in zuigelingen (5,4-6,3).
De blindedarm levert een zuurstofarme kamer voor de groei van bacteriën als Bacteroides spp., waarvan er wel 109/g aanwezig zijn. In combinatie met ingenomen voedsel dienen mucopolysacchariden van de bekercellen in de slijmlaag als een belangrijke bron van suikers voor de vertering van voedsel door de Bacteroides spp. Een rapport beschrijft een grote anaerobe bacterie als het voornaamste organisme aanwezig in de blindedarm van 108 tot 1010/ mL. Coliform bacteriën worden maar zelden aangetroffen in de blindedarm van konijnen. Verwacht wordt dat ze in kleine getale aanwezig zijn onder normale omstandigheden, maar drastisch in hoeveelheid toenemen als er ook maar een kleine toename in pH in de blindedarm (door ziekte of overlijden) optreedt.Daarom is een bacteriekweek zelfs van pure Escherischia coli niet noodzakelijk diagnostisch voor E. coli gerelateerde diarree. Andere criteria als epidemiologisch bewijs en histopathologische bevindingen moeten ook overwogen worden. Ook verschillende andere organismen komen voor in de blindedarm, namelijk Bifidobacterium spp., Endophorus spp., Streptococcus spp., en Acuformis spp. in de holte, Clostridium, Peptococcus, Peptostreptococcus, en Fusobacterium soorten aan het slijmvlies en allerlei ongeïdentificeerde niet-zuurstof afhankelijke soorten. Veel onschadelijke protozoën komen voor in de blindedarm, zoals gecilieerde protozoën gelijkend Isotricha van knaagdieren (107/mL), flagellate protozoën als Eutrichomastix spp., Enteromonas spp., en Retortamonas spp., en een amoeboid organisme, Entamoeba cuniculi.
Een gist die specifiek is voor konijnen, Saccharomyces (syn. Cyniclomyces) guttulatulus is aanwezig in 106/g en wordt vaak gevonden in uitstrijkjes van uitwerpselen. Lactobacilli zijn opvallend genoeg afwezig in de normale konijnendarm. Het gebruik van Lactobacilli als probiotica voor zieke konijnen komt vaak voor. Hoewel Lactobacillus acidophilus zeer goed in staat is om de zuurgraad van de maag te overleven, is de bruikbaarheid van deze bacterie discutabel.
Naar ervaring van de schrijver zijn enkele waterafhankelijke Lactobacillus producten voordelig voor sommige dieren, maar het effect is mogelijk eerder toe te wijzen aan de veranderingen in inname van electrolyten en vloeistoffen dan aan bacteriële kolonisatie.
De gecombineerde microbiële flora in de blindedarm verzorgt de afbraak van ammonium, ureum, eiwitten en enzymen van de dunne darm en cellulose (bij voorkeur in deze volgorde. Deze microben kunnen ook xylan en pectine metaboliseren. De producten van deze reacties zijn de eiwit- en enzymstructuren van de microben zelf (die later verteerd worden als keutels) en bijproducten van microbiële vertering, voornamelijk bestaande uit vrije vetzuren (acetisch, formisch, propionisch en butyrische zuren). Deze vrije vetzuren worden actief opgenomen door de wanden van de blindedarm en dikke darm en gebruikt als energiebron, net als bij andere knaagdieren. Konijnen verschillen van andere dieren in het opzicht dat de butyrische vetzuurconcentratie hoger is dan de concentratie van propionische vetzuren. De verhoudingen van vetzuren in de blindedarm zijn normaalgesproken 60-70% acetisch, 15-20% butyrisch en 10-15% propionisch zuur. Bij toename van de hoeveelheid vezels in de voeding en langere tijd van niet-eten wordt de hoeveelheid acetisch zuur verhoogd in de blindedarm. Gesuggereerd wordt dat butyrisch zuur mogelijk een remmend effect heeft op de peristaltiek en dus kan de verlaging van butyrisch zuur een verklaring zijn voor het feit dat toename van ingenome vezels leidt tot verhoogde darmactiviteit. Vrije vetzuren worden in vergelijkbare proporties aangetroffen in het bloed, wat suggereert dat vrije vetzuren voor het grootste deel onveranderd worden opgenomen. Desalniettemin duidt de aanwezigheid van lactaat in het bloed op een gering parietale stofwisseling.
Preventie van het eten van keutels leidt niet tot veranderingen van vrije vetzuren in het bloed, maar hoge concentratie vrije vetzuren in het spijsverteringskanaal zorgt voor een verhoging van het eten van keutels.
De dikke darm en fusus coli: anatomie en fysiologie
De dikke darm van het konijn is te verdelen in een aantal verschillende compartimenten op positioneel en morfologisch gebied. Het opstijgende deel van de dikke darm is erg lang en onderverdeeld in vijf delen, zich heen en weer uitstrekkend, gescheiden door buigingen. Het eerste deel beschikt over drie delen die drie rijen van zakjes vormen, ook wel ‘haustrae’ genoemd. In het tweede en derde deel zijn de delen gecombineerd tot een rij of haustra. De rest van het opstijgende deel van de dikke darm beschikt niet over deze delen (tenae), en ligt ingesloten in het achterste deel van de buikholte. Blindedarmkeutels kunnen voor het eerst duidelijk worden onderscheiden bij het einde van het opstijgende deel van de dikke darm.
Het transversale (dwarse) deel van de dikke darm is maar kort en eindigt in een verdikking, de fusus coli genaamd, een unieke structuur voor konijnen, hazen en dergelijke. De scheiding van de dikke darm in twee anatomische en fysiologische delen heeft geleid tot het gebruik van proximaal (dichtbij) en distaal (ver weg) in plaats van het originele gebruik van opstijgend, transversaal en dalend. De fusus coli is een soort pacemaker voor de peristaltiekgolf in de proximale en distale dikke darm en reguleert de scheiding van verteerbaar en niet-verteerbaar materiaal door samentrekkingen van de tenae/haustrae.
Wetenschappers hebben samenhang aangetoond tussen de activiteit van de fusus coli en verschillende regulatiesystemen, waaronder invloeden van het autonome zenuwstelsel en circulerende hoeveelheden prostaglandines, aldosteron en andere stoffen. Tijdens productie van harde keutels is de hoeveelheid aldosteron het hoogst. Het is echter niet bekend of dit de oorzaak of het gevolg is van de activiteit van de fusus coli.
Van prostaglandines is aangetoond dat ze een remmend effect hebben op de beweeglijkheid van het proximale deel van de dikke darm en de activiteit in de distale dikke darm stimuleren, daarbij de productie van blindedarmkeutels stimulerend.
De betrokkenheid van het autonome zenuwstelsel en de adrenaline klieren in de regulatie van de fusus coli zijn mogelijk de oorzaak van het feit dat konijn gevoelig zijn voor stress-gerelateerde ziekten aan het spijsverteringsstelsel. Na de fusus coli krijgen het afdalende deel van de dikke darm en het rectum hun simpele buisachtige vorm met dikkere wanden weer terug.
Voedselinname en het eten van blindedarmkeutels: tijdsafhankelijke relativiteit
Blindedarmkeutels zijn samenklonteringen van gedeeltelijk verteerde voedingsstoffen uit de blindedarm en dus geen onverteerde vezels (in normale keutels) en worden gevormd in de anus. Omdat het hier geen afvalstoffen betreft, is het strict genomen geen feces, hoewel blindedarmkeutels ook wel zachte feces genoemd worden. Het eten van blindedarmkeutels wordt foutief ook wel eens onder een noemer met coprophagia (het eten van eigen feces) genoemd. De blindedarmkeutels worden onder invloed van een neurologische lik-reflex direct gegeten wanneer ze uit het rectum worden uitgescheiden en worden in hun geheel doorgeslikt. Het eten van blindedarmkeutels (cecotrophie) wordt beïnvloed door het licht en het eetpatroon en verschilt tussen tamme en wilde konijnen. Cecotrophie bij wilde konijnen vindt vooral overdag plaats wanneer ze in hun holen zitten. Dit staat in contrast met tamme konijnen, waarbij cecotrophy voor het grootste deel ‘s nachts plaatsvindt, maar het kan ook op ieder moment van de dag plaatsvinden. Wanneer cecotrophy plaatsvindt, staat de overige voedselinname even stil en omgekeerd. Dit mechanisme kan niet worden verklaard door de vulling van de maag of voldoening, omdat het aanbrengen van een halsband die cecotrophy voorkomt geen veranderingen teweegbrengt in de formatie van blindedarmkeutels en ook het voedselpatroon niet verandert. Het hongergevoel wordt beïnvloed door verschillende factoren, waaronder een droge mond en verminderde bloedwaarden van metabolieten als glucose, aminozuren en vrije vetzuren. Cecotrophy volgt normaal gesproken zo’n vier uur na de inname van voedsel. Konijnen in gevangenschap in een normaal dag- en nachtritme en met een eindeloze voorraad voedsel, vertonen verhoogde voedselinname tussen 15.00 en 17.00 uur en voedselinname blijft hoog tot middernacht. Tot 2.00 uur ‘s nachts is er sprake van een verminderde voedselinname, waarna weer een verhoging optreedt die piekt rond 6.00 uur ‘s morgens en eindigt rond 8.00 uur. Cecotrophy treedt dus vooral op tussen 0.00 en 2.00 uur ’s nachts en weer om 8.00 uur ‘s morgens. Wanneer konijnen volgens een strak schema gevoed worden, is cecotrophy direct gerelateerd aan de inname van het voedsel en niet meer aan het daglicht. De mate van cecotrophy is direct gerelateerd aan de hoeveelheid vezels in de voeding. Cecotrophy is het hoogst bij een hoog percentage niet-verteerbare vezels. Vezels zijn volgens een vaste verhouding verdeeld tussen harde keutels en blindedarmkeutels. Blindedarmkeutels bevatten ongeveer 50% van de vezels van harde keutels en dit is onafhankelijk van de hoogte van vezelinname. Wanneer de inname van vezels echter wordt beperkt, daalt de hoeveelheid vezels in harde keutels drastisch, terwijl het vezelgehalte van blindedarmkeutels ongeveer gelijk blijft.
Beweeglijkheid van de blindedarm en de dikke darm en de formatie van blindedarmkeutels
Het belangrijkste proces van de spijsvertering van het konijn is de regulering van de beweeglijkheid van de blindedarm en de dikke darm om de scheiding van niet-verteerbaar afval en bruikbare verteerbare substraten mogelijk te maken. Het proces wordt gereguleerd door de beweeglijkheid van de dikke darm en kan ruwweg worden verdeeld in twee fases: de harde feces fase en de zachte feces fase (formatie van blindedarmkeutels).
De harde feces fase
Aanvankelijk is de blindedarm relatief leeg, aangezien de inhoud is verwijderd in de laatste zachte feces fase. Materiaal uit de dunne darm uit het ingenomen voedsel gaat door de ileocecale klep en sacculus rotundus en wordt in gelijke mate verdeeld over de blindedarm en het eerste deel van de dikke darm. Slechts weinig vertering vindt in deze fase plaats, de blindedarm trekt samen, waardoor het grootste deel van de inhoud in de dikke darm terechtkomt. Door de wanden van de dikke darm wordt water toegevoegd, dat de menging en scheiding bevordert. Onder stricte controle van de fusus coli vinden drie verschillende samentrekkingen plaats. Er is een progressieve monofasische peristaltische golf van vijf seconden en een segmentale lage-frequentie, 14 seconde durende samentrekking. Deze gaan beide in de richting van de mond naar de anus. De derde samentrekking is die van de haustra, die hoge-frequentie, drie seconde durende samentrekkingen ondergaan, die herhaaldelijk de inhoud van de dikke darm mengen. De stromingen van vloeistoffen en vaste deeltjes die zo ontstaan, zorgen ervoor dat onverteerbare vezels van groter dan 0,5 mm zich verzamelen in het midden van het eerste deel van de dikke darm. Kleinere deeltjes worden meer naar de zijkanten verplaatst, waardoor ze in de zakvormige haustra terechtkomen. De centrale vezelrijke massa wordt snel verder verplaatst en wordt omgevormd in harde feces door de samenpersende acties van de fusus coli. Absorptie van water, electrolyten en vrije vetzuren vindt plaats wanneer de feces verder door de dikke darm verplaatst wordt en uiteindelijk wordt de harde feces uitgescheiden als kleine, droge, harde keutels. Harde keutels hebben geen slijmlaag.
Wel-verteerbare materialen en vloeistoffen die zich hebben verzameld in de haustra, worden door terugwaartse peristaltiek teruggestuwd naar de blindedarm voor vertering.
Formatie van blindedarmkeutels – de zachte feces fase
Na vertering vormen de materialen in de blindedarm een zachte donkergroene pasta, die rijk is aan half-verteerde materialen en micro-organismen. Verschillende mechanismen die mogelijk de formatie van blindedarmkeutels aansturen, zijn reeds gesuggereerd, waaronder toegenomen concentraties van vrije vetzuren, eetgedrag en de aanwezigheid van voedsel in de maag en dunne darm in combinatie met de controlerende processen voor fusus coli-functie, die reeds zijn besproken. De samentrekkingen die nodig waren voor de scheiding van verteerbare en niet-verteerbare materialen nemen nu af. Peristaltische, monofasische samentrekkingen vinden nu elke 1,5 seconde plaats. De blindedarm trekt samen en de inhoud wordt snel voortgestuwd naar de dikke darm. De passagetijd van zachte feces door de dikke darm is 1,5-2,5 keer korter dan die van harde feces.
De samentrekkingen van de fusus coli gedurende de productie van blindedarmkeutels zijn soepeler en behouden het vochtgehalte van de darminhoud. Bekercellen in de fusus coli scheiden slijm uit. Wanneer de blindedarmkeutels de dikke darm passeren wordt lysozyme toegevoegd en worden de samenklonteringen voorzien van een slijmlaag. De blindedarm-keutels komen aan bij de anus en worden direct na uitscheiding ingenomen onder invloed van allerlei factoren, waaronder stimulatie van mechanoreceptors, olfactorische prikkels en bloedwaarden van verschillende metabolieten en hormonen.
Samenvatting
Het spijsverteringskanaal van het konijn is zeer complex, dat zich concentreert op de scheiding van verteerbare en niet-verteerbare componenten van het voedsel in het eerste deel van de dikke darm. Belangrijk voor dit systeem is de aanwezigheid van vezels met lange stam ( >0,5 mm) om de beweeglijkheid van de blindedarm en de dikke darm te waarborgen. Het grootste deel van de problemen met de spijsvertering van tamme konijnen zijn te wijten aan slechte voeding (weinig vezels; veel eiwit; veel suikers) en het voeren van tussendoortjes op niet-vaste tijden, waaraan het konijn niet gewend is. Veel van deze problemen kunnen voorkomen worden wanneer tamme konijnen een dieet krijgen dat voornamelijk bestaat uit vezels, als gras, hooi en kruiden. Het voeden van fruit, granen en snoepjes met veel suiker of vet kan beter worden vermeden. Samengestelde voedingen kunnen het beste, hoewel ze handig zijn, minimaal gegeven worden. Wanneer er toch voor gekozen wordt om dit voedsel te verschaffen, kan het beste gekozen worden voor de voedingen waarbij extrusie is toegepast, waardoor de lange vezels behouden blijven.
Fig. 1. Schematische weergave van het spijsverteringsstelsel van het konijn. (Uit: Harcourt-Brown F. Textbook of rabbit medicine. Oxford: Butterworth-Heinemann; 2002)
spijsvertering
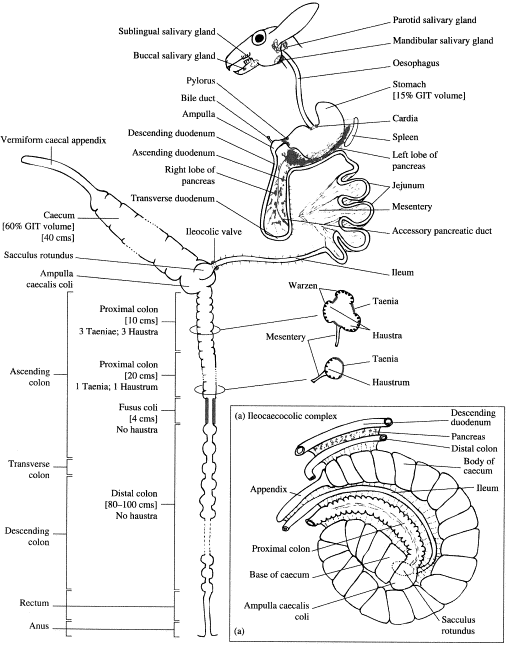
Fig. 2. Een weergave van de activiteit van het verteringsstelsel van het konijn. (Overgenomen van Harcourt-Brown F. Textbook of rabbit medicine. Oxford: Butterworth-Heinemann; 2002)
Vertaald uit het Engels door: CJ Masson
Bron:
2003 Elsevier Science (USA).